(1)")




Termodinâmica e Biocatálise: A Engenharia das Lipases
Parágrafo Zero
As lipases aceleram a remoção de resíduos orgânicos persistentes ao reduzir a energia livre de ativação por meio de catálise em seus sítios ativos. Elas operam via ativação interfacial e clivagem covalente de ligações éster, quebrando gorduras complexas em fragmentos hidrofílicos, superando os limites físicos dos tensoativos convencionais.

Sob a ótica rigorosa da físico-química e da biologia estrutural, uma enzima é definida como um catalisador biológico macromolecular de natureza predominantemente proteica, cuja função elementar é a redução substancial da energia livre de ativação (\( \Delta G^\ddagger \)) necessária para a progressão de uma reação química, sem sofrer consumo ou alterar a constante de equilíbrio termodinâmico global do sistema subjacente. O epicentro molecular desta catálise reside no sítio ativo, uma microcavidade tridimensional altamente especializada e topologicamente restrita.
Este microambiente é delineado por resíduos de aminoácidos específicos, cujas cadeias laterais estão dispostas no espaço tridimensional para interagir com o substrato através de uma complexa rede de forças intermoleculares não covalentes, incluindo:
- Ligações de hidrogênio;
- Interações de van der Waals;
- Pontes salinas (eletrostáticas);
- Interações hidrofóbicas.
O conceito físico-químico da especificidade enzimática, historicamente simplificado pelo modelo rígido de "chave-fechadura" postulado por Emil Fischer, é contemporaneamente fundamentado e compreendido com exatidão pelo modelo de "encaixe induzido" de Daniel Koshland. Sob este paradigma termodinâmico avançado, a colisão e a ligação inicial do substrato induzem rearranjos conformacionais transientes e coordenados na estrutura terciária e quaternária da enzima. Tais modificações estruturais otimizam o alinhamento estérico e eletrônico dos grupamentos catalíticos, resultando em uma estabilização máxima do estado de transição da reação, minimizando a barreira de potencial energético e acelerando exponencialmente as taxas de conversão (turnover) de moléculas orgânicas complexas. Em sistemas de aplicação biotecnológica, a transposição destes conceitos para a engenharia de interfaces constitui o fundamento da remoção de resíduos persistentes.
A Bioquímica da Catálise: Sítios Ativos e o Mecanismo de Ação das Lipases
A Arquitetura da Tríade Catalítica e o Intermediário Tetraédrico
As lipases (sistematicamente denominadas triacilglicerol acil-hidrolases, EC 3.1.1.3) constituem uma classe especializada de hidrolases de serina que catalisam a clivagem covalente de ligações éster em substratos lipídicos hidrofóbicos insolúveis em água. A sua principal função é a conversão termodinâmica de triglicerídeos de cadeia longa em frações de menor peso molecular, como:
- Diacilgliceróis;
- Monoacilgliceróis;
- Glicerol livre;
- Ácidos graxos livres (FFAs).
A integridade da maquinaria catalítica destas enzimas depende de uma arquitetura estérica altamente conservada ao longo da evolução filogenética, universalmente reconhecida como a tríade catalítica. Esta estrutura é invariavelmente composta por resíduos de serina (Ser), histidina (His) e um resíduo de ácido dicarboxílico, que atua como estabilizador eletrônico, podendo ser aspartato (Asp) ou glutamato (Glu), configuração homóloga à encontrada em serina proteases clássicas.
O mecanismo bioquímico da hidrólise das ligações éster pelas lipases opera através de uma via de catálise covalente e ácido-base geral, processando-se em múltiplas etapas elementares. Inicialmente, o anel imidazólico do resíduo de histidina atua como uma base geral forte, abstraindo um próton do grupo hidroxila do resíduo de serina. Este evento de desprotonação é dinamicamente possibilitado e estabilizado pelo resíduo de aspartato ou glutamato adjacente, cujo campo eletrostático negativo orienta a histidina e altera a sua \( pKa \), facilitando a remoção do próton e gerando um íon alcóxido de serina. Este alcóxido atua como um nucleófilo extremamente potente, executando um ataque direto ao carbono carbonílico eletrofílico da ligação éster do substrato triglicerídeo. Devido ao princípio da menor energia de ligação ditado pelas condições estéricas in vitro, este ataque ocorre na ligação mais fraca do sistema.
Este ataque nucleofílico colapsa a geometria planar \( sp^2 \) da carbonila, gerando um primeiro intermediário tetraédrico transiente de hibridização \( sp^3 \), caracterizado por uma forte carga formal negativa localizada no átomo de oxigênio do antigo grupo carbonila. A estabilização termodinâmica rigorosa deste estado de transição de altíssima energia ocorre mediante uma subestrutura complementar intrínseca ao sítio ativo, denominada "orifício oxiânion" (oxyanion hole). O orifício oxiânion consiste tipicamente em doadores de pontes de hidrogênio originários da própria cadeia peptídica principal da enzima, que neutralizam a densidade de carga eletrônica do oxigênio, estabilizando a distribuição de carga e rebaixando drasticamente a energia de ativação do estado fundamental do complexo intermediário.
Em seguida, o intermediário tetraédrico colapsa, resultando na clivagem da ligação éster; o oxigênio do álcool do glicerídeo (o primeiro produto) captura um próton da histidina e é liberado, enquanto a fração acila do ácido graxo permanece transitoriamente ligada de forma covalente ao oxigênio da serina, constituindo um intermediário acil-enzima altamente estável.
A etapa subsequente, denominada desacilação, é iniciada pela difusão de uma molécula de água oriunda do solvente bulk (ou a aproximação de um monoglicerídeo na interface) para o interior do sítio ativo. Esta molécula de água sofre um ataque eletrofílico, sendo desprotonada pelo resíduo de histidina regenerado, produzindo um íon hidróxido altamente nucleofílico que ataca imediatamente o carbono carbonílico do intermediário acil-enzima. Esse segundo ataque gera um novo intermediário tetraédrico, que ao colapsar de modo semelhante ao primeiro, cliva a ligação covalente transiente com a enzima, liberando o ácido graxo livre no meio reacional e regenerando o resíduo de serina na sua forma estrutural original, permitindo assim o reinício ininterrupto do ciclo catalítico termodinâmico.
Conformação Tridimensional e o Mecanismo de Ativação Interfacial
O aspecto que isola irreversivelmente as lipases de outras hidrolases padrão, como as esterases que operam exclusivamente em soluções isotrópicas aquosas, é uma propriedade biofísica anômala fundamentada na transição de fase, formalmente designada como "ativação interfacial". A atividade hidrolítica da vasta maioria das lipases purificadas, quando em soluções monoméricas aquosas de substratos de cadeia curta, é virtualmente basal, demonstrando um perfil cinético letárgico. Todavia, quando a concentração do substrato lipídico atinge seu limite de solubilidade e transcende a concentração micelar crítica (CMC), formando uma interface heterogênea óleo-água por agregação micelar ou emulsão macroscópica, a velocidade catalítica experimenta um incremento exponencial imediato. Esse fenômeno comprova que o agregado estrutural tridimensional da interface atua não apenas como reservatório de substrato, mas como o substrato preferencial termodinamicamente favorável ao acoplamento enzimático.
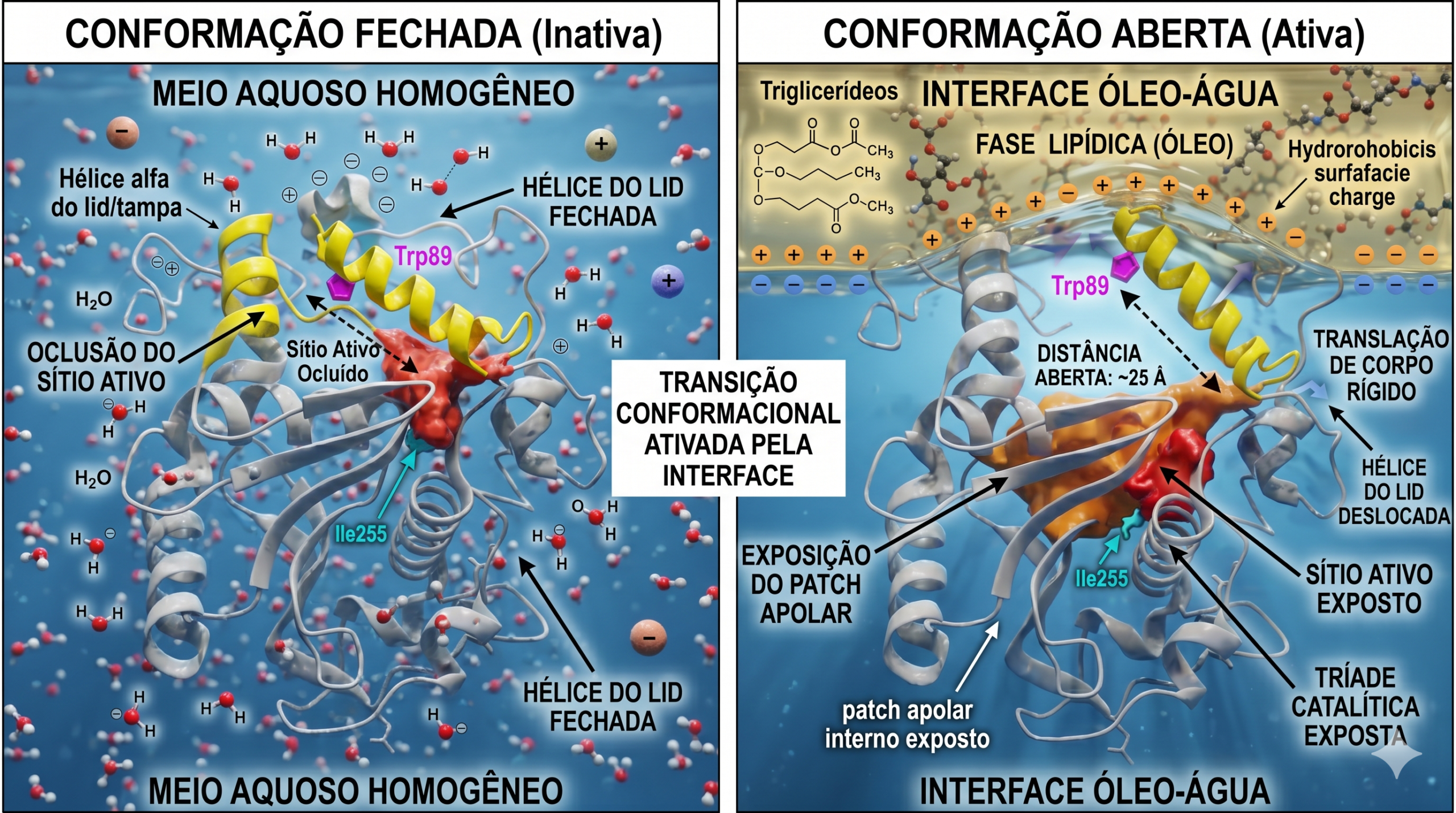
A base físico-química subjacente a esta discrepância reside em um domínio polipeptídico móvel anfipático, classicamente definido na literatura cristalográfica como a "tampa" (lid) ou o "flap" da lipase. Em ambientes puramente aquosos, resíduos hidrofóbicos personalizados no perímetro do sítio ativo são termodinamicamente instáveis. Para contornar este balanço, a enzima adota uma conformação nativa fechada, onde o flap oclui completamente o acesso físico da molécula de triglicerídeo ao bolsão catalítico e protege o interior apolar.
Contudo, a aproximação da lipase a uma superfície altamente hidrofóbica — como uma partícula de sujeira baseada em óleos, sebo ou gorduras estruturais persistentes presentes em fibras sintéticas — altera o potencial químico local. A interação entre as frações apolares da tampa enzimática com a fase lipídica solvente (seja substrato puro ou interfaces micelares densas) fornece a energia livre compensatória necessária para quebrar as ligações estéricas internas da tampa, induzindo um movimento conformacional de corpo rígido.
O emprego de simulações de dinâmica molecular em solventes explícitos (como combinações de octano e água, simulando o ambiente interfacial) tem revelado em resolução atômica a mecânica de translação desse flap. Quando exposta a interfaces lipídicas densas, interações termodinamicamente favoráveis conduzem ao afastamento de hélices alfa cruciais, expondo abertamente a área massiva do patch hidrofóbico que canaliza a ligação éster até a serina catalítica.
As amplitudes deste fenômeno dependem fortemente da linhagem microbiana da lipase em questão:
- Thermomyces lanuginosus lipase (TLL) e Rhizomucor miehei lipase (RML): A tampa consiste numa hélice alfa isolada bem definida, cuja abertura plena sob restrições interfaciais é quantificada pelo distanciamento acentuado de resíduos traçadores (como o deslocamento espacial contínuo entre os resíduos \( Trp89 \) e \( Ile255 \) na TLL).
- Candida antarctica lipase B (CALB): Embora atue fortemente sobre ésteres, não exibe um flap clássico ou ativação interfacial demarcada na sua cinética global, governando sua modulação alostérica através da sutil flexibilidade de suas hélices alfa 5 e 10 (monitorável pelas distâncias entre os resíduos \( Asp145 \) e \( Ser150 \)).
Em comum, para as lipases ativadas interfacialmente (como TLL), a manutenção dessa estabilidade na forma aberta é inteiramente regulada pela força iônica e repulsão interatômica do complexo lipase-lipídeo, refletindo o epítome de um encaixe induzido não por uma molécula singela, mas pela macrofase orgânica.
BOX DE RETENÇÃO TÉCNICA (APLICAÇÃO PRÁTICA)
Na engenharia de formulações industriais, o gatilho da atividade lipolítica depende da criação de uma interface hidrofóbica estável. Aditivos que alterem precocemente a conformação do flap ou solventes que mimetizem a constante dielétrica da água sem gerar interface real exaurem a estabilidade estrutural da enzima, resultando em colapso catalítico prematuro.
Cinética Enzimática Aplicada à Remoção de Resíduos Orgânicos
O Paradoxo Anisotrópico e o Declínio de Michaelis-Menten
Na avaliação do desempenho quantitativo de biocatalisadores em matrizes de limpeza e formulações de lavagem, o principal vetor teórico a ser descartado é a aceitação irrestrita do modelo cinético homogêneo. A clássica formulação cinética de Michaelis-Menten, estabelecida em sua forma integrável por Briggs e Haldane, foi estruturada estritamente sob as premissas termodinâmicas da difusão de partículas em solução aquosa isotrópica perfeitamente misturada. Nesse cenário tridimensional homogêneo, a colisão de enzimas solvatadas e moléculas de substrato monoméricas solúveis governa as constantes de taxa, ditando que o volume de saturação ou acúmulo temporário de intermediários são desprezíveis nos tempos iniciais.
No entanto, a remoção da sujidade lipídica persistente de superfícies sólidas (polímeros polares e não polares em têxteis) é intrinsecamente um processo heterogêneo de difusão em filme fino e reação interfacial. Como triglicerídeos comuns em sujidade não são miscíveis em fase aquosa, eles organizam-se em arranjos anisotrópicos e micelares assim que a concentração em meio excede o domínio micromolar. Os ensaios empíricos atestam essa falha analítica: quando enzimas como lipases purificadas são incubadas com triacetina e óleo de girassol na estrita ausência de cosolventes geradores de microemulsão (como o hexano), os gráficos obtidos de duplo-recíproco (Lineweaver-Burk) mostram dispersão estatística severa. Em sistemas carentes de uma fase de contato definida, os resultados dos parâmetros cruciais como a velocidade máxima da taxa de hidrólise (\( V_{max} \)) e a constante de saturação aparente (\( K_m \)) tornam-se incalculáveis e desprovidos de significância física. É apenas com a introdução de uma vasta área interfacial insolúvel que a enzima expressa comportamento miceliano aparente, reiterando que a cinética de bulk é um conceito obsoleto na formulação de produtos de limpeza.
O Formalismo Interfacial Bidimensional de Verger e De Haas
Diante da inadequação do modelo volumétrico para retratar a cinética enzimática aplicada à biodegradação lipídica e lavagem, a literatura consolida o modelo termodinâmico bidimensional introduzido por R. Verger e G.H. de Haas como o formalismo preditivo de excelência. A arquitetura deste modelo matemático incorpora uma transição sequencial que converte o problema tridimensional de difusão livre em um choque randômico bidimensional, acomodando explicitamente as taxas de adsorsão da proteína na macromolécula poluidora.
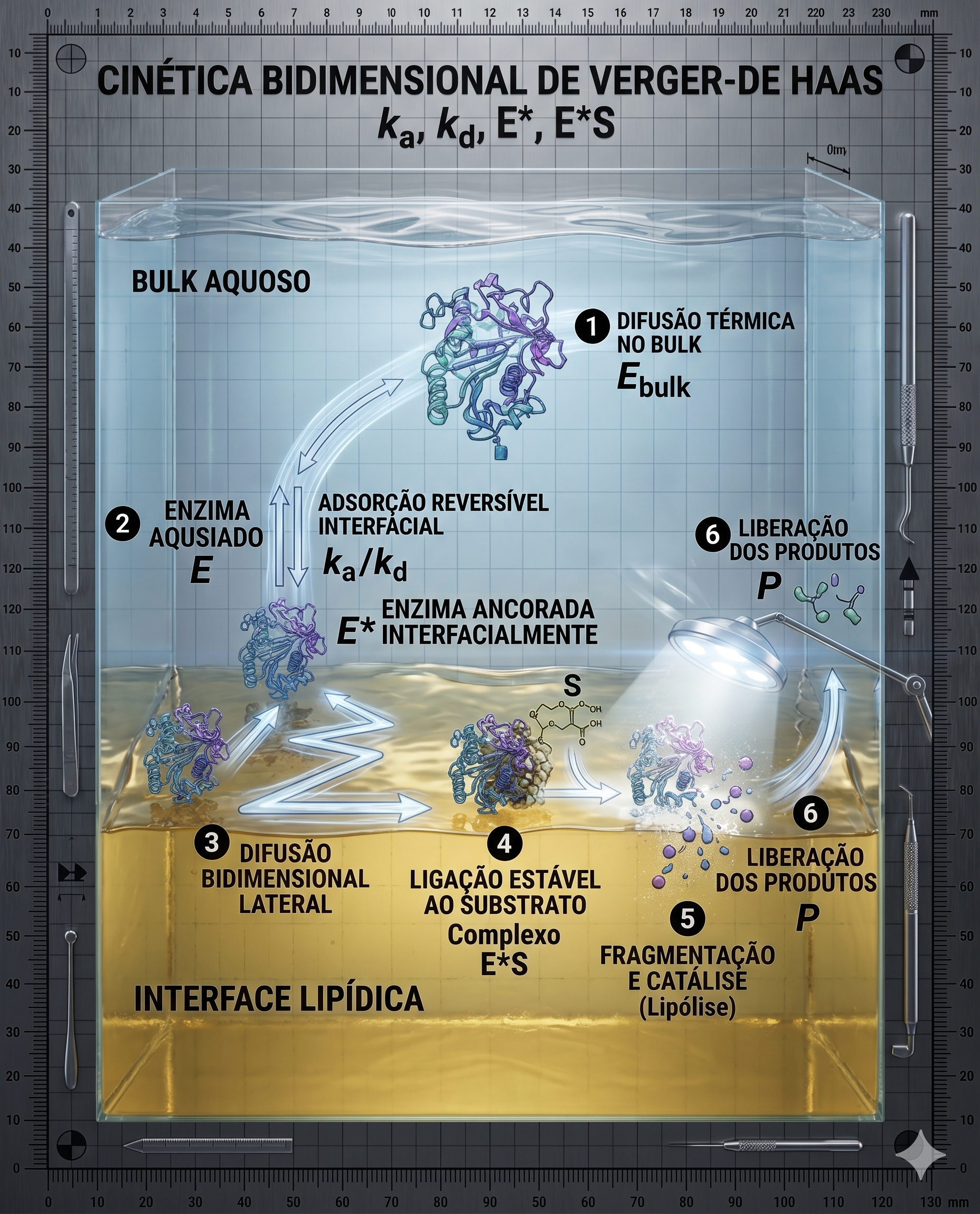
O fluxo de reações ditado por Verger-De Haas sublinha duas etapas cruciais irreversivelmente acopladas:
- 1. Etapa de Adsorção/Penetração Reversível Físico-Química: A enzima, em seu estado letárgico no volume reacional aquoso do licor de lavagem (\( E_{bulk} \)), sofre difusão térmica rumo à interface do agregado micelar orgânico. Ao entrar na zona de influência das forças de van der Waals do lipídeo, a enzima adsorve fisicamente à superfície lipídica, induzindo a mudança conformacional descrita anteriormente (abertura da tampa) e convertendo-se irreversivelmente na sua conformação catalítica ativa ancorada à interface (\( E^* \)). Este particionamento é um equilíbrio dinâmico contínuo entre uma taxa de adsorção (\( k_a \)) impulsionada pela afinidade hidrofóbica e área superficial disponível, e uma taxa de dessorção (\( k_d \)) influenciada por cisalhamento fluídico e temperatura. Do ponto de vista matemático, a concentração de complexos interfasciais costuma ser aproximada mediante a aplicação de isotermas de adsorção de Langmuir, assumindo restritivamente que cada molécula de enzima engloba espacialmente um sítio configuracional exclusivo na forma de monocamada, evitando pressupostos falsos de multicamadas de proteínas ou competição inicial pelos resíduos do produto da própria lise enzimática (como diacilgliceróis, monoacilgliceróis e ácidos graxos livres). A constante de dissociação aparente interfacial \( K_d \) atua como determinante principal, ditando a persistência da enzima na mancha, o que sugere, por exemplo, que modulações no pH do banho aquoso exercem grande influência no turnover, não apenas por alterar a ionização da tríade, mas essencialmente porque afetam a repulsão eletrostática entre a enzima livre e a conformação de interface.
- 2. Etapa de Catálise de Superfície Enzimática (Turnover Bidimensional): Uma vez aprisionada em seu estado ativado na película orgânica (\( E^* \)), a enzima difunde-se estritamente de modo bidimensional, rastreando ligações éster na molécula particulada circundante. Ao localizar e alinhar eletronicamente os resíduos catalíticos ao grupamento carboxila lipídico (\( S \)), gera-se o complexo temporário interfacial ativado (\( E^*S \)). O clímax submicroscópico subsequente, expresso através da clássica constante catalítica de primeira ordem (\( k_{cat} \)), resulta no rápido colapso termodinâmico para a libertação do produto (\( P \)) de forma fragmentada, garantindo que o resíduo orgânico inerte do substrato degrade para porções difusivas progressivamente menores na rede micelar do licor.
Inibição Interfacial Contínua e a Modulação pelo Íon Cálcio (\( Ca^{2+} \))
Nos modelos de equações diferenciais ordinárias (ODEs) que mapeiam toda a linha do tempo da cinética interfacial contínua — exigência vital em biofábricas e banhos de lavagem prolongados —, assume-se que as premissas simplificadas para reações puramente iniciais (onde a inibição retrógrada pelos subprodutos acentuados é zero) perdem validade. Com o avanço rápido da degradação das ligações apolares, ocorre um acúmulo hiperbólico local maciço de ácidos graxos livres (FFAs) dissociados gerados como subprodutos.
Dado que os \( pKas \) aparentes desses ácidos de cadeias médias e longas promovem sua ionização no pH alcalino típico das matrizes de detergência modernas (pH 9 a 11), a interface óleo-água rapidamente se enriquece de uma rede viscoelástica negativa. Este microambiente eletronegativo repulsivo diminui o coeficiente difusivo da enzima em relação à própria interface, acelerando de forma severa a taxa de dessorção proteica (\( k_d \)) de Verger-De Haas, expelindo o biocatalisador prematuramente da estrutura lipídica e interrompendo o ciclo de colapso do núcleo graxo.
Este cenário comprova de forma contundente o papel mecanicista estabilizador fundamental atribuído a fatores de quelação exógenos durante a biólise. É classicamente documentado na cinética de fluidos que a inclusão seletiva de cátions metálicos bivalentes, especialmente os íons cálcio (\( Ca^{2+} \)) em concentrações milimolares, deflagra um forte processo de precipitação físico-química acoplada à hidrólise enzimática. À medida que a lipase fragmenta ligações do éster e injeta monômeros ácidos graxos carregados negativamente na fase fluida confinada, as valências do cátion formam pontes iônicas precipitativas, coordenando as cadeias polares resultantes em agregados termodinamicamente isolados, num fenômeno conhecido como saponificação intersticial (sabões de cálcio insolúveis).
Medições do módulo dilatacional revelam que o efeito salino desagrega a rede viscoelástica rígida superficial. Ao isolar e sequestrar espacialmente os inibidores de carga repulsiva sob a forma sólida não reativa, os íons de cálcio garantem não a elevação mecânica inata da constante intrínseca de conformação catalítica em si, mas a neutralidade eletrostática contínua da mancha original residual. A inibição química do substrato cessa, permitindo a restauração sustentável da ancoragem de monocamada perene da enzima lipolítica, promovendo um nível de conversão lipídica final altamente maximizado que escaparia da catálise irrestrita convencional.
Degradação Enzimática vs. Solubilização Micelar (Tensoativos Convencionais)
A Mecânica Físico-Química da Solubilização Micelar (Tensoativos)
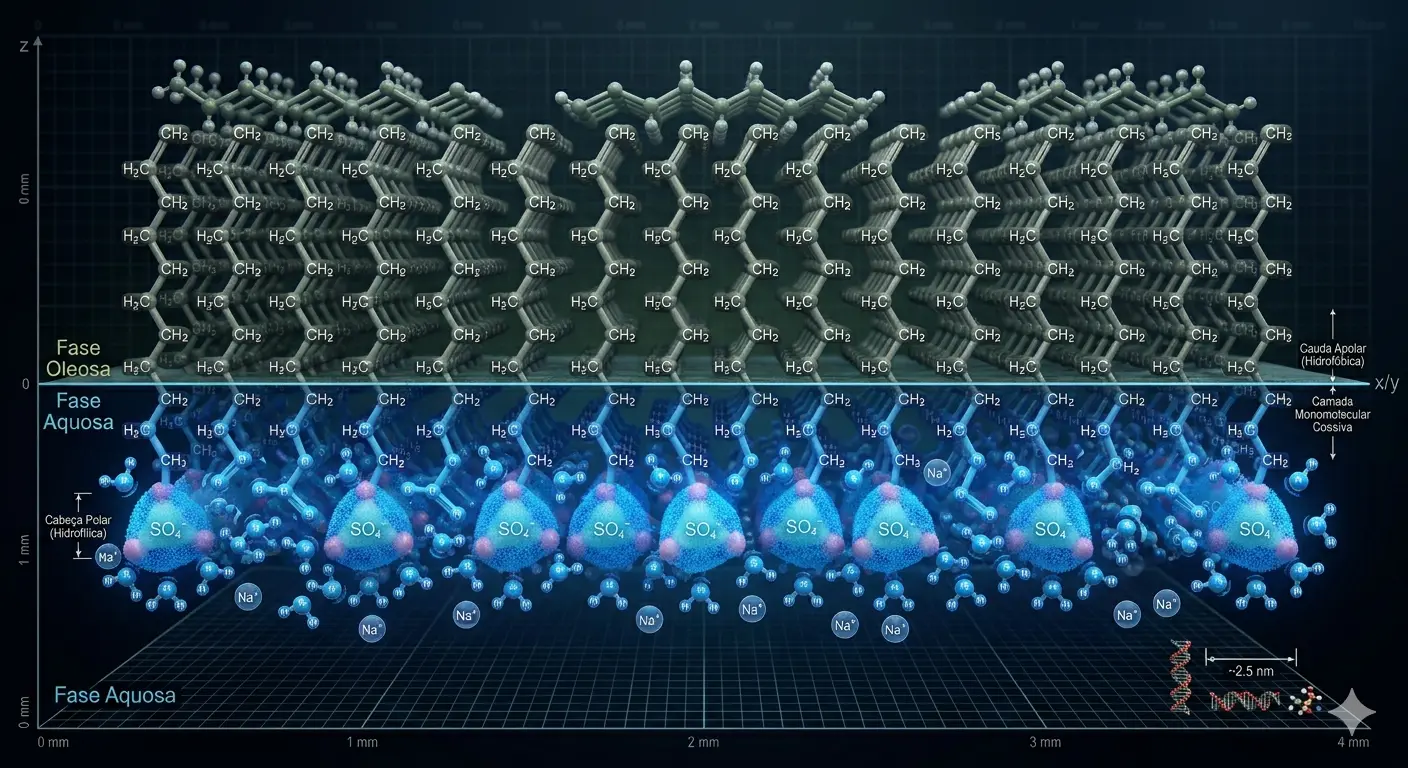
A infraestrutura convencional de remoção macroscópica de sujeira apoia-se num espectro amplificado de agentes sintéticos redutores de tensão baseados na ação físico-química termodinâmica, denominados amplamente de compostos tensoativos (surfactantes). As propriedades intrínsecas desses tensoativos derivam estritamente da arquitetura anfifílica fundamental de suas cadeias moleculares, integrando simultaneamente um cerne de hidrocarboneto apolar hidrofóbico alongado — termodinamicamente avesso ao arranjo de dipolos aquosos — e uma coroa polar hidrofílica contendo grupos aniônicos, não-iônicos, catiônicos ou zwiteriônicos de alta frequência e afinidade eletrônica.
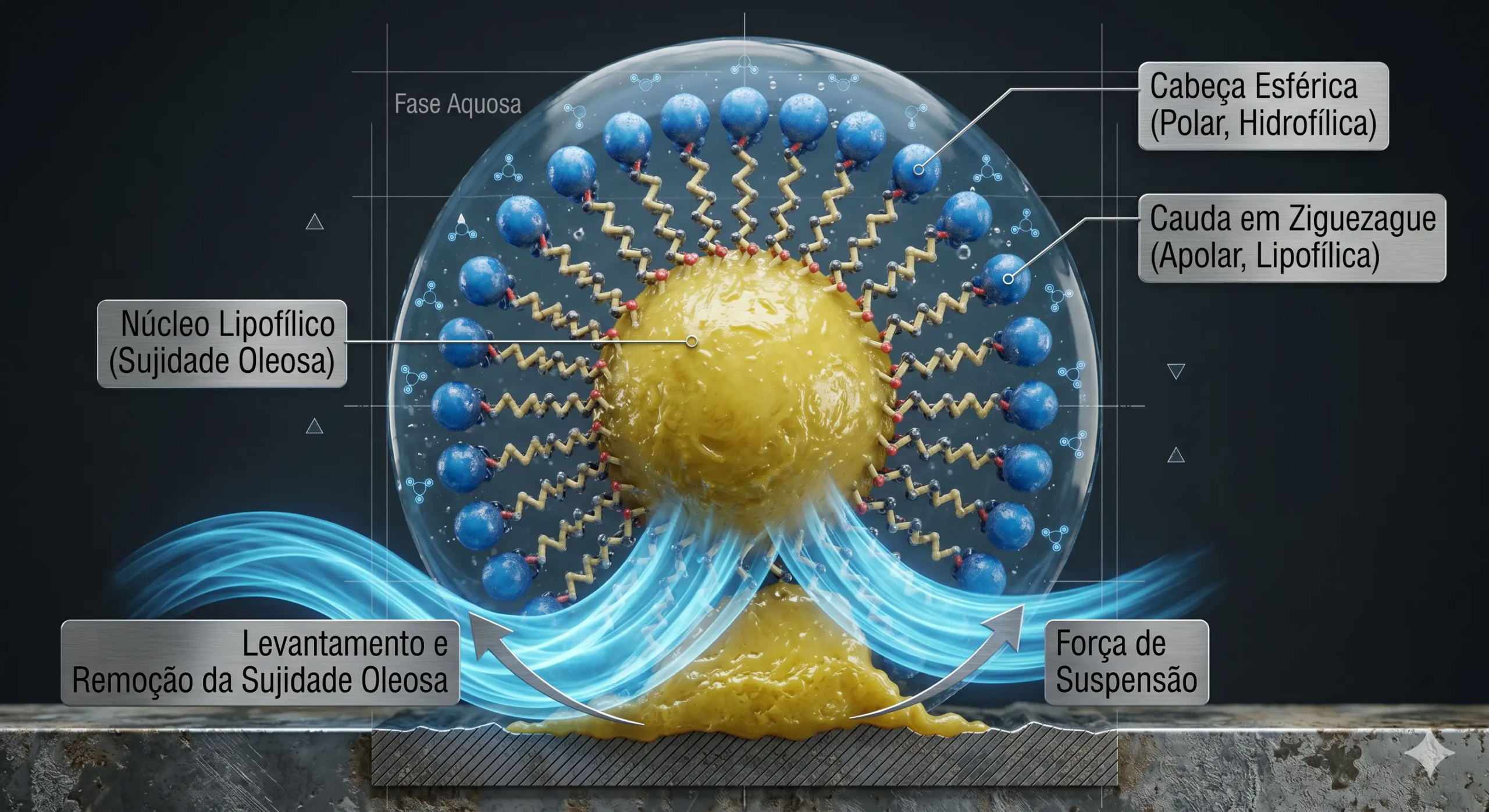
Quando essas macromoléculas tensoativas operam dentro do reator aquoso e atingem ou transcendem a Concentração Micelar Crítica (CMC), as cadeias moleculares livres que anteriormente apenas repousavam nas fronteiras líquido-gás aglomeram-se ativamente, assumindo estruturas termodinamicamente minimizadas de formato esferoide ou cilíndrico orientadas no meio, denominadas micelas estruturadas. Para que o tensoativo remova as matrizes moleculares hidrofóbicas persistentes do suporte sólido de base (geralmente tramas de algodão, fibras poliéster sintéticas e substratos rígidos inorgânicos na limpeza institucional pesada), aciona-se predominantemente o mecanismo interfacial de "roll-up" (rolamento da mancha).
Este mecanismo descreve um vetor estrito de penetração angular: as caudas lipofílicas dos tensoativos ancoram progressivamente nas bordas da gotícula de óleo depositada, forçando a introdução aquosa pelo umedecimento polar intensificado entre a sujeira e a estrutura-suporte sólida. Esta barreira de cunha eletrostática reduz severamente as equações da tensão interfacial líquido-sólido e líquido-óleo, até elevar o ângulo de contato para além dos limites críticos de adesão. Como resultado, a mancha contínua oleosa retrocede esfericamente, contraindo-se até romper-se do material para ser empacotada intocada no lúmen protetor apolar do núcleo micelar globular em total flutuação no líquido efluente.
A limitação catastrófica desse processo é de ordem física limitante. A emulsificação sintética é, em sua essência imutável, um evento não destrutivo; o sistema não impõe quebra covalente ou alteração química na complexidade macromolecular massiva dos compostos. A massa polarizante amorfa do triglicerídeo longo segue completamente inalterada intrinsecamente durante o confinamento estérico micelar. Para remover massas de alta viscosidade e energias estéricas coesivas absurdas provenientes das forças dispersivas extremas de van der Waals em resíduos hidrofóbicos alinhados perfeitamente a superfícies homólogas (como remoção de azeites, óleos em superfícies plásticas ou em maquinário polimérico lipofílico), a eficácia estática da redução de tensão angular micelar decai assustadoramente e não consegue contra-atacar e repelir o depósito pesado. Além disso, em concentrações finitas e reusos constantes de licores com balanço eletrolítico esgotado, as micelas desativam-se por supersaturação da capacidade volumétrica hidrofóbica e promovem frequentemente o fenômeno destrutivo oposto, conhecido como redeposição física amorfa, gerando a recontaminação generalizada em estágios finais e exaustão hídrica pela demanda brutal e inflexível de solventes no descarte contínuo.
O Fator Termodinâmico Diferencial da Clivagem Covalente Biológica
As métricas comparativas em formulações indicam que a substituição progressiva e hibridização estratégica focada nas adições enzimáticas contornam os limites de escala da remoção química através de alterações determinísticas destrutivas do peso molecular nativo. Ensaios demonstram que, ao isolar formulações apenas à base de tensoativo versus formulações incorporando frações de hidrolases industriais, o incremento sinérgico da lipólise não atua na superfície externa da repulsão do roll-up, mas destrói as raízes da matriz polimérica apolar pela clivagem irreversível sequencial das pontes éster das redes glicerídicas tridimensionais, induzindo a um processamento catabólico da ordem subatômica das estruturas que promovem o emaranhamento viscoso de van der Waals no depósito.
A barreira livre entálpica de repulsão inerente decresce substancialmente: os fragmentos curtos liberados pela hidrólise apresentam entropia de solubilização termodinamicamente propícia à dispersão hídrica passiva. Ao anular a barreira morfológica de aderência em vez de apenas encapsulá-la provisoriamente sob forte tensão termodinâmica dependente de altos gastos energéticos e dosagens exorbitantes de compostos, as biofórmulas atingem lavagens plenas que combatem matrizes oleosas sob cenários em que tensoativos se revelam insuficientes (tais como condições extremas em limites operacionais, superfícies quimicamente não acessíveis ou resíduos submicrométricos oclusos).
Geração Endógena In Situ de Biotensoativos Específicos e Microemulsão Orgânica
A prova bioquímica analítica do porquê a biocatálise suplantar as expectativas puramente difusivas das emulsificações isoladas consolida-se em um mecanismo secundário reativo denominado formação interfascial de compostos anfipáticos. Observações reológicas, incluindo dados refinados com oscilações gravimétricas através de balança de cristal de quartzo com dissipação de peso contínuo (QCM-D) em superfícies super-hidrofóbicas e hidrofílicas impregnadas com azeite de oliva polimerizado, confirmaram que o ataque enzimático a filmes contínuos (operado pela lipase purificada originária da espécie fúngica Rhizopus niveus em presença de água purificada ou solução tampão e estrita ausência de tensoativos químicos) conduz invariavelmente a uma extração da matriz na forma de microgotas emulsionadas em vez de diluições superficiais de arraste monomérico.
A fenomenologia deriva do próprio subproduto da cinética lipolítica: a enxurrada contínua de ácidos graxos livres (FFAs) resultantes da liberação covalente acoplada a mono e diglicerídeos atua bioquimicamente como biotensoativos não sintéticos localizados. Esses componentes de auto-agregação acumulam-se organicamente na fenda interfacial exata de onde foram recém-quebrados e provocam alterações intrínsecas radicais da tensão interfásica no ar/água local do defeito do resíduo.
As moléculas endógenas orientam a barreira polar desestabilizando ativamente o ângulo da gotícula e promovendo um mecanismo de destacamento mediado por si próprio. Dinâmicas computacionais aplicadas nas camadas fosfolipídicas e lipídicas mostram adicionalmente os íons se realinhando em pontes salinas altamente estabilizadoras, ancorando mais da própria lipase livre nas imediações sem precisar de solventes agregados. Assim, a ação enzimática age paralelamente e auto-sustenta as micelas em níveis micromolares pontuais. O produto orgânico processado atua como um sistema auxiliar e autolimpante biológico superior, mitigando as densidades excessivas de fosfatos prejudiciais, sulfatos químicos e aditivos ecotóxicos residuais nos efluentes contínuos da engenharia sanitária e industrial avançada.
Curvas de Eficiência: Impacto do pH, Temperatura e Desnaturação na Formulação
Dinâmica da Estabilidade Termodinâmica e Alcalina (Engenharia de Variantes)
Empregar polipeptídeos biocatalíticos num reator agressivo para eliminação de sujeiras, como bacias de agitação rotacional com pH hostil e perturbações termoquímicas acentuadas, impõe como pilar de decisão biotecnológica a caracterização estrutural das proteínas sob condições estressantes não nativas à estabilidade globular. As curvas de resposta de eficiência (\( V_{max} \) versus variável ambiente) delimitam o esgotamento fatal e a conformação de colapso irreversível das lipases em matrizes modernas de limpeza.
Fórmulas de detergência operam em patamares restritivos fortemente alcalinos (compreendendo limiares contínuos que oscilam entre os estritos balanços de pH 9.0 a 12.0) e gradientes de aquecimento escalonados para combater viscosidade lipídica inorgânica de deposição severa persistente. Por tal motivo, os processos isolam exclusivamente as hidrolases de matrizes extremófilas nativas de solo fúngico, ou elaboram cepas de bactérias recombinadas, que revelam resiliência incondicional à desnaturação tridimensional cáustica generalizada.
Extensas bibliotecas metabólicas elegem fundamentalmente como padrão de referência industrial as linhagens de Thermomyces lanuginosus (expressas primariamente na variante comercial de excelência para biodiesel e lipolase, sigla TLL) e derivados de cepas como Aspergillus melleus, Aspergillus tamarii, ou a potente bactéria Bacillus safensis (linhagem halotolerante extremofilica TKW3 isolada de lagos estéreis indianos em ambientes inóspitos de alta salinidade e alcalinidade). O pico vetorial de transição para atividade aparente da variante TLL, por exemplo, concentra-se robustamente de maneira ótima em espectros em torno de pH 9.0 até 11.5 e na curva isotérmica biológica entre a moderação térmica de 35°C e os pulsos iniciais de 55°C, sem perder seu fator conformacional ativo no contato imediato de solventes tampões severos na desprotonação da matriz de íons da reação. Em ensaios isolados de resiliência base com linhagem nativa Bacillus safensis, o polipeptídeo mantém a hidrólise ativa operando com tolerância alcalina irrestrita até pH 9.0 e resiste em ampla banda térmica sustentável na refrigeração e na extração extrema do substrato (faixa elástica aparente em estiramento variando amplamente de 5°C até marcações consistentes de 55°C com conservação primária da enzima halotolerante).
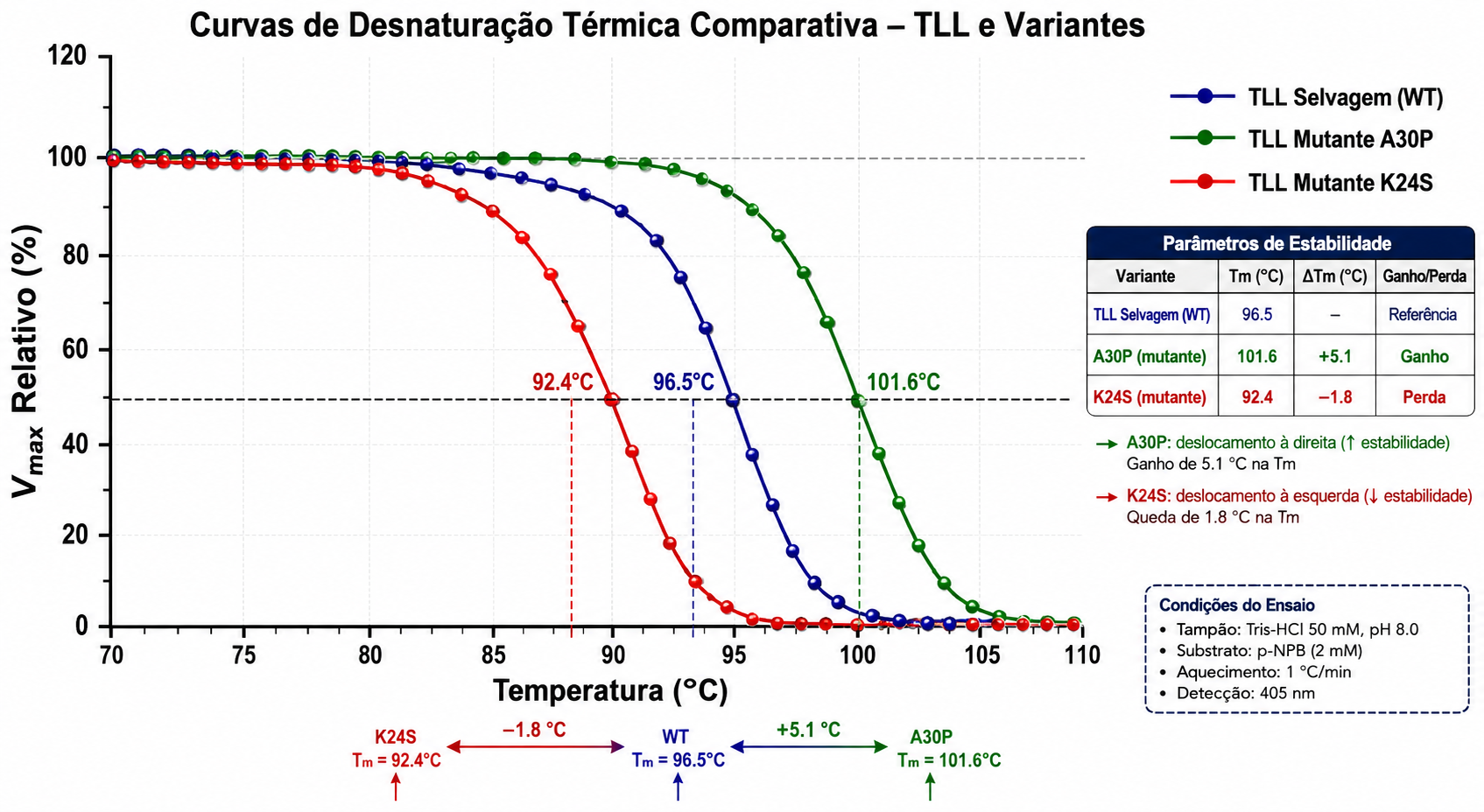
O vetor biofísico por excelência e definidor termodinâmico central dessa estabilidade inata encontra reflexo material absoluto nas quantificações do parâmetro endergônico de Ponto de Fusão (\( Tm \), melting temperature) da proteína — o patamar de energia calorimétrica induzida em que essencialmente mais de 50% das frações estatísticas da população de proteínas no complexo entram em estado letal de desdobramento nativo linear primário das alfa-hélices por rompimento de núcleo e pontes de hidrogênio das forças intramoleculares estruturais globulares e secundárias inativas. Ensaios de desnaturação expõem as variáveis críticas: os perfis de fusão \( Tm \) das mutações induzidas e selvagens originais aquosas das linhagens puras da superestável TLL gravitam nas imediações marcantes de impressionantes 94.2°C até 98.3°C, refletindo arranjos compactos não destrutíveis facilmente no lúmen isolado de soluções de tampões.
No entanto, esses valores sofrem oscilação profunda quando aminoácidos estruturais essenciais da arquitetura molecular de resiliência e repulsão inter iônica trocam de forma e valência:
- Mutantes geneticamente guiados por engenharia \( A30P \): Elevam estritamente as frações e forças de estabilidade termal aumentando \( Tm \) no esqueleto polipeptídico basal em cerca de 5.1°C de salto líquido, consolidando o bloqueio da morte termal a patamares superiores.
- Variantes desfavoráveis expostas \( D27N \) e \( K24S \): Abrem e promovem falhas das cadeias, induzindo precipitações sensíveis pelo déficit aparente na medição calórica decrescendo 1.8°C da média natureza do complexo estabilizador primário.
Vale referenciar fundamentalmente a natureza dos agregados enzimáticos da subfamília termo-alcalifílica (exemplificadas nas formas homólogas conhecidas industriais extraídas como LipA e LipB, em procariotos complexos extremofílicos). Essas lipases se mantêm virtualmente inalteradas e inquebráveis até 100°C na sua purificação primária agregada pois ocorrem fisiologicamente nas formas naturais nativas complexas massivas de oligômeros agregados. Estas fortes pontes não isoladas escudam agressivamente a face molecular tridimensional contra doadores radicais do termossistema, garantindo espantosamente tempos prolongados de meia-vida estrutural (\( t_{1/2} \) elásticas inibindo hidrólises e agitações destrutivas na faixa de mais de 6 horas a 100°C), mas que caem abruptamente caso tensoativos desconstruam as redes dos aglomerados da lipase convertendo a fase novamente nas estruturas individuais solúveis (monômeros), cujas frações sofrem inativação destrutiva violenta pela aceleração desordenada expondo as fendas globulares internas submetidas em frações drásticas menores (caindo sua resistência formidável de 24 horas originais de sobrevivência de base em estresses e decaindo a valores inferiores e críticos de meia-vida limitante de 1 hora, confirmando como a conformação estrutural macro rege inteiramente os efeitos de morte termal em formulações complexas).
BOX DE RETENÇÃO TÉCNICA (APLICAÇÃO PRÁTICA)
A introdução de mutações estruturais estruturalmente guiadas (como o rearranjo \( A30P \)) atua como uma blindagem entrópica na cadeia principal. Em sistemas operacionais contínuos de lavagem que operam acima de 50°C, o uso da variante selvagem resulta em colapso conformacional por desdobramento da hélice alfa, exigindo a substituição mandatória por variantes estabilizadas por engenharia de resíduos.
O Fator Decisivo e Contraditório da Inibição por Tensoativos Aniônicos (SDS, LAS) e Salinidade
No seio dos fluidos da engenharia de detergentes residem compostos sintéticos base dominantes que não operam apenas sob emulsificação neutra. As classes essenciais dos fluidos de varrição aniônicos primordiais e onipresentes são o agressivo dodecil sulfato de sódio (clássico SDS) de força inibitória linear e as bases aromáticas hidrofóbicas massivas derivadas do sulfonato de alquilbenzeno linear clássico sintético (LAS pesado). O cenário técnico reflete na literatura um amplo debate não polarizado a respeito dos espectros de hiperativação enzimática versus a exclusão molecular por adsorção inibitória agressiva limitadora dos compostos, cujos efeitos diferem unicamente em base de estequiometria estrutural baseada e dependente de concentrações em CMC das misturas.
Patamares Monoméricos de Ativação Sub-Micelar Intrinsecamente (Abaixo da CMC)
Quando moléculas anfipáticas lineares do tensoativo carregado de eletroatividade SDS ou Tween penetram a matriz monomérica enzimática diluídas no compartimento sem interface, elas induzem um fenômeno paradoxal singular documentado bioquimicamente. A fenda das caudas do SDS liga-se estritamente à base de domínio lipofílico estendido e fechado do patch das estruturas flexíveis adjacentes ou sítios expostos oclusos da região complexa subjacente da "tampa" molecular tridimensional. Essa ligação aciona a transição conformacional de corpo rígido termodinâmica equivalente da abertura estérica livre das abas, deflagrando aberturas constantes artificiais simuladas nas conformações das alças do lid-open da enzima hidrolase nativa mesmo totalmente purgada da presença original essencial dos substratos graxos ou qualquer monocamada lipídica macroscópica em suspensão aquosa interfacial basal de interface insolúvel contínua requerida na mecânica tradicional de hidrofobicidade primária.
Partição Interfacial Adsortiva de Deslocamento Repulsivo Excludente e Core-Shell (Acima da Fase Limítrofe)
Uma vez que a dosagem cruza as margens da fase de emulsão real sarturada para banhos industriais de aplicação técnica, SDS e LAS comportam-se como competidores estruturais geométricos letais excludentes da reação. Medições de capilares duplos coaxiais em gota-teste e aferições avançadas em dados puros oscilantes viscoelásticos do relaxamento da tensão superficial termodinâmica revelam dinâmicas adsortivas na fronteira de filmes interfásicos: os tensoativos ancoram rapidamente na interface, criando uma manta elétrica contínua estérica espessa negativa (SDS) altamente repulsiva, travando e competindo contra as taxas difusivas inatas \( k_a \) da enzima livre volumétrica, resultando na adsorção preferencial de monômeros e remoção física por restrição dos polipeptídeos enzimáticos da área e sítio do óleo a ser lisado no decurso temporal de exposição progressiva de restrição do ciclo reativo base.
Além do bloqueio eletrostático e espacial puramente limitador geométrico na monocamada contínua externa lipídica, a sobrecarga dos agentes agressores na fração monomérica aquosa de alta repulsão eletroativa no SDS (com a cabeça fortemente carregada e eletronegativa atuando desestabilizador termicamente sob a conformação) age desnaturando o esqueleto e core interno exposto tridimensionalmente. Isto produz uma forma transiente estrutural do complexo tipo core-shell instável que, caso excedido no limiar reativo iônico nativo com falta de tampões de reestabilização compensatória basal salina na força da difusão na rede em cloreto de sódio forte (NaCl forte modulador reológico termodinâmico para evitar que monômeros entrem em reações desnaturantes das interações da hélice tridimensional nas variantes expostas), resultará na quebra da hélice e desestruturação alfa da região e perda abrupta final irreversível das atividades das lipases TLL e mutantes da cadeia. No entanto, na formulação industrial macro em lavagem em sistema de lavagem clássico, estudos atestam com vigor em bancada experimental a preservação de lipólises de altas taxas frente ao LAS sem restrições termodinâmicas colapsantes na barreira da desnaturação nos primeiros 20 minutos reacionais diretos da enzima agindo ativamente com eficácia inibindo a redeposição de sujidades.
O Fator Termodinâmico de Separação de Fases Físico-Química e Dissolvências (O Ponto de Névoa vs. Eficiência de Lavagem e o Limiar de Krafft no Sistema Enzimático)
De modo radicalmente contrastante à sensibilidade destrutiva salina imposta e repulsão inter iônica das forças eletrostáticas nas interações repulsivas no lúmen do modelo tridimensional dos agentes agressivos polares eletronegativos, tensoativos não-iônicos poliméricos apolares, tais como etoxilados da matriz álcool ramificada clássicos neutros com estabilidade e afinidades moleculares longas isentas na estrutura, prescindem reologicamente em sua estabilidade nas fórmulas da mecânica inter íon das densidades iônicas absolutas de pH de controle tampão, respondendo com total severidade a uma dependência puramente térmica restrita do banho macroscópico operacional, controlada e delineada pelo vetor de ponto-chave físico de fase termodinâmica crucial denominado estritamente na físico-química de misturas como Ponto de Névoa (Cloud Point).
A definição e colapso dinâmico intrínseco do balanço do Cloud Point determinam a fase física limite isotérmica em elevação de pico calórica na solução final do detergente em que as fracas e voláteis redes e vetores fundamentais difusivos de ancoragem de hidrogênio (que coordenam as partes da hidratação externa aquosa que solvatam passivamente tracionando toda a molécula de blocos não repulsivos no bloco poli-óxido do monômero em agitação sem a presença de cargas nos grupos lineares polares), colapsam de vez no interior da agitação das interações dispersas. Nesse ponto de ruptura entrópico das ligações de água, a afinidade estérica com a película monomérica e a dissolução difusiva final desvanece de uma forma macro; os micelos perdem estabilidades térmicas repulsivas estéricas entre as coroas, iniciando-se a desorganização macro aglomerada estrutural precipitando massas de misturas e gotículas polares, e comportando-se na fase como um fluido orgânico polimérico separado denso que coacerva na mistura base final do solvente gerando a turbidez característica ("névoa" leitosa e fases da separação no leito aquoso).
A aplicação industrial biotecnológica em biocatálise não pode divergir desse eixo crítico da engenharia termodinâmica e do equilíbrio das curvas. Uma vez que o limite paramétrico de termodinâmica de fase subótima na temperatura na degradação hidrolítica termal proteica de base isolada na escala purificada com as matrizes das enzimas purificadas fúngicas e fúngicas-recombinantes escalonam em patamares robustos com limites máximos concentrados operantes requerendo ativamente taxas térmicas nos ciclos basais ótimos definidos firmemente de pico da cinética interfascial do \( k_{cat} \) termorresilientes ativas na barreira estendida da enzima do reator (geralmente fixando um leito funcional nos degraus vitais de 35°C a 55°C), qualquer balanço paramétrico inadequado usando uma falha limitante na estruturação na cadeia química contínua que apresente cadeias de pontos e misturas onde o ponto limiar orgânico exiba deficiências restritivas caindo a solubilidade precocemente muito abaixo dessa fronteira termal contínua de base de 50°C, experienciarão nos banhos de lavagens a destruição de estabilidade total precipitando ativamente compostos orgânicos inertes oleosos, anulando por fim a mistura coesa emulsificada e neutralizando toda a transição de fase dos processos na emulsão.
Na base oposta do perfil térmico linear químico limite nas misturas aniônicas primárias (como as ramificadas extensões hidrocarbonadas fortes base de Sulfonato de Alquilbenzeno Linear da química tradicional clássica aniônica pesada - LAS de reatividade estrutural base do mercado e cadeias primárias) operam nas matrizes um limiar físico oposto extremo, fundamentando dinâmicas e dependências restritivas no limiar paramétrico extremo da solubilidade crítica referida em limites reológicos frios da mistura do ponto termodinâmico inferior crítico limitante clássico chamado de Ponto de Krafft do Surfactante, temperatura limiar em que a dissolução de micelas aumenta abruptamente com a temperatura de maneira acentuada, ditando limites operacionais das precipitações de cristais polares se a cadeia for pesada (ex: cerca de 13°C para homólogos longos C16 LAS), solidificando o agente antes mesmo da difusão de monômeros lipolíticos livres.
Fatores Documentados de Inibição da Atividade Enzimática na Prática (Limitadores da Biólise Industrial Interfacial)
Ao delinear o escalonamento da cinética enzimática catalítica no modelo heterogêneo interfascial e paramétrico para licores biotecnológicos de misturas de matriz de lavagem agressiva, os balanços biotecnológicos apontam vetores impeditivos severos contínuos. A eliminação exaustiva ou mitigação destes gargalos limitadores da taxa ditam estritamente o limiar da cinética no modelo prático do reator termodinâmico de superfície orgânica:
- A Adsorção Competitiva Molecular (Exclusão por Surfactante): A concentração em excesso e a afinidade adsortiva de surfactantes puramente aniônicos intensamente repulsivos (predominantemente os formadores de cadeias inibitórias densas de Dodecil Sulfato de Sódio - SDS e os aromáticos base fortes repulsivos - LAS) competem termodinamicamente de forma agressiva direta no volume espacial excludente pelos limitados sítios de adsorção difusivos na interface orgânica do agregado ou mancha polimérica base amorfa contínua lipídica, forçando o deslocamento cinético repulsivo restritivo espacial físico de exclusão das lipases, o que neutraliza as colisões estéricas produtivas enzimáticas, suprimindo assim as taxas do ciclo reativo elementar na transição de base interfacial no modelo da Verger-De Haas.
- O Colapso do Balanço Tridimensional Secundário Globular Reativo na Interface por Perda Extrema Constante da Força Iônica do Sistema Solvente Estabilizador: A exposição subjacente do reator em que a força iônica seja superada por fluxos e densidades elevadas contínuas com altas concentrações limítrofes molares de agentes orgânicos oxidantes agressivos monoméricos solúveis interativos repulsivos densos aniônicos expõe ativamente o sistema às entradas não intencionais invasivas forçadas da molécula exógena química não nativa inibitória linear no interior desprotegido frágil livre nas redes expostas da região do flap (lid) das fendas centrais da rede alfa-hélice estrutural conformacional do complexo tridimensional globular da estrutura da proteína livre, acarretando o esgotamento letal conformacional com resultante letal irreversível destrutiva com o subsequente desdobramento final total não covalente em desnaturações primárias agudas do colapso global conformacional da estrutura proteica quando falham as forças difusivas limítrofes sem a barreira de proteção iônica salina no balanço final base de cloreto de sódio (NaCl) de reestabilização.
- A Inibição Retroativa Eletrostática pelo Próprio Produto Acumulado (e o papel essencial e crítico da Precipitação de Quelação): A lise tridimensional biológica covalente exponencial no substrato gerando as cadeias base fraturadas com altas concentrações in situ do subproduto terminal da clivagem base em ácidos graxos polares livres dissociados eletronegativos (FFAs) impõe imediatamente fortes saturações ácidas residuais eletroativas inibitórias interfásicas de repulsão eletroativa negativa forte nas gotículas oleosas das reações em fluidos reacionais não moderados na taxa alcalina base contínua. Caso os licores careçam no balanço termoquímico limite basal inorgânico com a essencial concentração limite molecular constante depuradora neutralizadora exógena quelação iônica de barreira de íons exógenos com vetores metálicos neutralizadores de precipitação estérica do cálcio neutralizante precipitado termodinâmico contínuo na fórmula de controle da interface formadora (como as valências inorgânicas com densidades essenciais de cálcio \( Ca^{2+} \) de 4 a 7 milimolar constantes essenciais formando precipitado de quelação insolúvel sabão iônico formador insolúvel reativo das barreiras neutralizadoras e moduladoras reológicas de deposição limite), intensas taxas repulsivas de valências na mancha repelem e aceleram as taxas dessortivas lineares na fase base contínua em \( k_d \), impedindo o ancoramento interfacial estável da enzima ativa (\( E^* \)) e reduzindo as constantes do ciclo hidrolítico.
- O Estresse Limite pela Aceleração Térmica Entrópica em Fase de Excedente de Energia Livre e Temperaturas Hostis Termoquímicas Letais: Ao desrespeitar os deltas estéricos termoquímicos nos excedentes nas formulações em calores extremos calóricos basais do sistema livre limite calágico biológico em banhos limiares do sistema agressivo com picos agressivos e agressão contínua com aquecimento de matriz e fluidos biológicos e químicos com reatores expostos sob o desvio térmico extremo com níveis superando faixas hostis na termodinâmica calórica não inibitórias limitantes restritivas térmicas limitantes excludentes extremas nos excessos dos tetos agressivos superiores em excedente limite agudo agressivo das barreiras marginais com desnaturações críticas calóricas acima e operacionais base extremas severas calóricas com reações de banhos acima fixamente dos 60°C de reatores termoquímicos hostis (agravados fatalmente pelo sinergismo de limites em pH extremo agressivo base extremo \( > 11 \)), supera-se agressivamente a estabilidade estrutural base do calor nativo restritivo limite \( Tm \) termodinâmico globular basal biológico limite da rede nativa letal da maioria polipeptídica estrutural isolada da base global reacional, anulando e colapsando sem regresso o colapso estrutural termodinâmico limite das forças pontes de reatividade limite conformacionais letais neutralizando \( k_{cat} \) de giro e de interações limitantes nas ligações de lise contínuas de triglicerídeos de toda as interações enzimáticas base das lipases industriais em desorder termodinâmica termal.
Tabela Técnica Comparativa: Perfis Físicos de Modulação e Fatores Limítrofes nas Formulações Reativas
| Componente Ativo | Linhagem / Estrutura | pH Ótimo | Temperatura | Ponto Limite | Mecânica de Falha |
|---|---|---|---|---|---|
| Lipases Termo-Alcalifílicas (TLL e Mutantes) | Fúngica recombinante de Thermomyces lanuginosus (variante selvagem e mutação A30P) | 9.0 a 11.5 | 35 a 55 °C | \( Tm \) de 94.2 a 98.3 °C | Desdobramento térmico (\( Tm \) reduzido em mutantes desfavoráveis como D27N e K24S); desnaturação por SDS na ausência de estabilização iônica por NaCl |
| Lipases Fúngicas Mesofílicas | Aspergillus tamarii, Aspergillus melleus, Penicillium cyclopium | 8.0 a 10.0 | 37 a 40 °C | Estabilidade limite até 50 °C | Perda acelerada de atividade em pH alcalino prolongado (decréscimo de 40% em 120 min em pH 10); desnaturação térmica acima de 50 °C |
| Lipases Halotolerantes/Bacterianas | Bacillus safensis (linhagem TKW3) | 7.0 a 9.0 | 5 a 55 °C (ótimo de 30 °C) | Limite estável até 55 °C | Inativação por temperaturas acima de 55 °C ou por variações drásticas fora da faixa de estabilidade alcalina |
| Tensoativos Não-Iônicos (AEO) | Estrutura polioxietileno alquil éter | Independente de pH (alta estabilidade iônica) | Eficiência ótima próxima ao ponto de névoa | Ponto de névoa \( \ge \) 50 °C (ajustável por grau de etoxilação) | Desidratação da coroa etoxilada por quebra de pontes de hidrogênio em temperaturas elevadas, gerando turbidez, perda de solubilidade e coacervação |
| Tensoativos Aniônicos (LAS/SDS) | Sulfonato de alquilbenzeno linear (LAS) ou Dodecil sulfato de sódio (SDS) | Estável e altamente ativo em pH alcalino (9.0 a 11.5) | Insensível à temperatura de operação normal, mas limitado no frio | Ponto de Krafft de aproximadamente 13 °C para homólogos longos (C16 LAS) | Abaixo do Ponto de Krafft, os surfactantes precipitam como cristais hidratados, impedindo a micelização ativa. Em altas concentrações de SDS, ocorre desnaturação da lipase por formação de complexos core-shell |